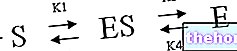Ver también: dieta cetogénica; cetoacidosis diabética.
Generalidad
En el pasado se pensaba que los cuerpos cetónicos se debían a un metabolismo excesivo, causado por la ingestión de demasiada grasa o diabetes. Los cuerpos cetónicos, por otro lado, son producidos naturalmente por nuestro cuerpo: el cerebro se adapta para usar estos metabolitos en condiciones de ayuno prolongado (en diabéticos, los cuerpos cetónicos reemplazan el metabolismo de la glucosa) Además, puede haber una exasperación de la vía de los cuerpos cetónicos en caso de mala nutrición.
Que son los cuerpos cetónicos
Los cuerpos cetónicos son derivados de los lípidos (derivan del metabolismo de los lípidos, casi exclusivamente hepático), pero tienen características que los hacen asemejarse a los azúcares:
- Alta velocidad de entrada;
- Rápido de usar.
Incluso algunos aminoácidos, en particular las condiciones metabólicas, pueden originar cuerpos cetónicos (leucina, lisina, fenilalanina, isoleucina, triptófano y tirosina).
Papel biológico
- Los cuerpos cetónicos son de pequeño tamaño, por lo que se transportan muy rápidamente (mucho más que los ácidos grasos que, por otro lado, necesitan proteínas de transporte como la albúmina);
- los cuerpos cetónicos son utilizados casi exclusivamente por los músculos y tejidos periféricos, pero también por el corazón (el 20-30% de la energía que utiliza proviene de los cuerpos cetónicos) y por el cerebro (en caso de ayuno prolongado).
Síntesis
Los cuerpos cetónicos son sintetizados por la acetil coenzima A, que se deriva del metabolismo de los ácidos grasos.
La enzima que cataliza la primera etapa es la Β-cetotiolasa, que aprovecha el azufre de la acetil coenzima A para producir una Β-ceto acil-coenzima A (es la reacción opuesta a la observada en la Β-oxidación de ácidos grasos); esta reacción no es espontánea, sino que es impulsada por la reacción posterior , catalizado de "hidroximetilglutaril coenzima A sintasa y que implica la unión de una segunda acetil coenzima A, obteniendo 3-hidroxi 3-metil glutaril coenzima A.
Posteriormente, interviene una enzima lítica que convierte la 3-hidroxi 3-metil glutaril coenzima A en acetato de vinagre, que es un cuerpo cetónico. El acetato de vinagre puede enviarse a los tejidos periféricos o, por la acción de la enzima hidroxi butirato deshidrogenasa, convertido en 3-Β-hidroxi butirato. Si el acetato de vinagre está en una concentración muy alta, también puede descarboxilarse espontáneamente a acetona.
Acetona, acetato de vinagre y 3-Β-hidroxi butirato son los tres cuerpos cetónicos que consideramos; la acetona es un producto de desecho, que se produce aleatoriamente en el camino de los cuerpos cetónicos y se expulsa por exhalación y transpiración.
Uso en tejidos periféricos
Los cuerpos cetónicos, producidos en el hígado, se envían a los tejidos periféricos.
Veamos, ahora qué sucede cuando el acetato de vinagre y el 3-Β-hidroxi butirato llegan a los tejidos periféricos. El acetato de vinagre es un Β-cetoácido, por lo tanto, si se activa, se puede utilizar en el proceso de Β-oxidación para producir acetil coenzima A: por lo tanto, es necesario transformar un Β-cetoácido en una Β-ceto acil coenzima A.
Cuando el acetato de vinagre llega a la mitocondria de una célula de un tejido periférico, se somete a la acción de la enzima. succinil coenzima A transferasa: a través de esta enzima, el acetato de vinagre reacciona con la succinil coenzima A (procedente del ciclo de krebs) y se obtienen succinato y vinagre acetil coenzima A.
Al explotar la succinil coenzima A, para activar el acetato de vinagre, saltamos al ciclo de krebs, la etapa que produce un GTP: este es el proceso, en términos de energía, que la célula está dispuesta a pagar para obtener la coenzima A de acetil vinagre. ; este último luego pasa bajo la acción de Β-ceto tiolasa (Enzima de oxidación Β) para producir dos moléculas de acetil coenzima A que se envían al ciclo de krebs.
Si el 3-Β-hidroxi butirato se envía a los tejidos periféricos, este último, dentro de la mitocondria, se convierte en vinagre acetona por la acción de la enzima Β-hidroxi butirato deshidrogenasa, con la producción de un NADH que corresponde a aproximadamente 2,5 ATP; el acetato de vinagre producido sigue el camino descrito anteriormente.
La célula de un tejido periférico extrae más energía del 3-Β-hidroxi butirato que del acetato de vinagre, pero el suministro de uno u otro a los tejidos periféricos depende de la disponibilidad de energía del hígado.
C "es una cantidad no despreciable de ácidos grasos metabolizados, contenidos en los peroxisomas y no en las mitocondrias; los peroxisomas son orgánulos más pequeños que las mitocondrias y ricos en iones metálicos y enzimas peroxidasa. Las enzimas peroxidasa usan peróxido de hidrógeno para promover los procesos redox, por lo tanto, en los peroxisomas hay es un sistema enzimático capaz de producir peróxido de hidrógeno.
En la Β-oxidación en peroxisomas, la "acil coenzima A, se obtiene por la acción de"acil coenzima A oxidasa (En la mitocondria, en cambio, actuó la enzima acil coenzima A deshidrogenasa.) También en este caso se forma la trans 2,3 enoil coenzima A, que sufre la acción de una enzima bifuncional (realiza la misma función que en las mitocondrias por "enoil coenzima A hidratasa y L-Β-hidroxi acil coenzima A deshidrogenasa) y así se convierte en Β-ceto acil coenzima A. Esta última, como en las mitocondrias, sufre la acción de Β-ceto tiolasa y acetil coenzima A y se obtiene una acil coenzima A con un esqueleto carbonoso reducido en dos unidades con respecto al de partida, que vuelve a la circulación.