Definición
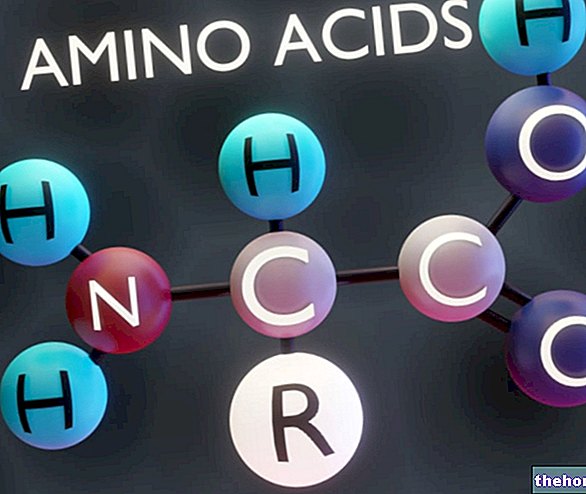
Las enzimas son proteínas producidas en células vegetales y animales, que actúan como catalizadores acelerando reacciones biológicas sin ser modificadas.
Las enzimas actúan combinándose con una sustancia específica para transformarla en una sustancia diferente; Los ejemplos clásicos los dan las enzimas digestivas presentes en la saliva, el estómago, el páncreas y el intestino delgado, que realizan una función esencial en la digestión y ayudan a descomponer los alimentos en constituyentes básicos, que luego pueden ser absorbidos y utilizados por el cuerpo, procesados por otras enzimas o excretado como desperdicio.
Cada enzima tiene una función específica: la que descompone las grasas, por ejemplo, no actúa sobre las proteínas ni sobre los carbohidratos. Las enzimas son esenciales para el bienestar del organismo. La deficiencia, incluso de una sola enzima, puede causar graves alteraciones. Un ejemplo bien conocido es la fenilcetonuria (PKU), una enfermedad caracterizada por la incapacidad de metabolizar un aminoácido esencial. fenilalanina, cuya acumulación puede provocar deformidades físicas y enfermedades mentales.
Estudio bioquímico
Las enzimas son proteínas particulares que tienen la característica de ser catalizadores biológicos, es decir, tienen la capacidad de descomponer la energía de activación (Eatt) de una reacción, modificando su camino para que un proceso cinéticamente lento sea más rápido.
Las enzimas aumentan la cinética de reacciones termodinámicamente posibles y, a diferencia de los catalizadores, son más o menos específicas: por tanto, poseen especificidad de sustrato.
La enzima no interviene en la estequiometría de la reacción: para que esto suceda, el sitio catalítico final debe ser idéntico al de partida.
En la acción catalítica casi siempre hay una fase lenta que determina la velocidad del proceso.
Cuando hablamos de enzimas no es correcto hablar de reacciones de equilibrio, hablamos, en cambio, de estado estable (estado en el que un determinado metabolito se forma y consume de forma continua, manteniendo su concentración casi constante en el tiempo). El producto de una reacción catalizada por una enzima suele ser en sí mismo un reactivo para una reacción posterior, catalizada por otra enzima, etc.
Los procesos catalizados por enzimas suelen constar de secuencias de reacciones.
Una reacción genérica catalizada por una enzima (E) se puede resumir de la siguiente manera:

E es la enzima
S es el sustrato;
ES representa el aducto entre la enzima y el sustrato;
P es el producto;
K es la constante de velocidad de la reacción.
Una enzima genérica (E) se combina con el sustrato (S) para formar el aducto (ES) con una constante de velocidad K1; puede disociarse de nuevo en E + S, con una constante de velocidad K2, o (si "vive" lo suficiente ) puede proceder a formar P con una constante de velocidad K3.
El producto (P) puede, a su vez, recombinarse con la enzima y reformar el aducto con constante de velocidad K4.
Cuando la enzima y el sustrato se mezclan, hay una fracción de tiempo en la que aún no se ha producido el encuentro entre las dos especies: es decir, hay un intervalo de tiempo extremadamente corto (que depende de la reacción) en el que la enzima y el sustrato han aún no se cumple; después de este período, la enzima y el sustrato entran en contacto en cantidades crecientes y se forma el aducto ES. Posteriormente, la enzima actúa sobre el sustrato y se libera el producto. Entonces se puede decir que c "es un intervalo de tiempo inicial en el que no se puede definir la concentración del aducto ES; después de este período, se asume que un estado estacionario Se establece, es decir, la velocidad de los procesos que conducen a la obtención del aducto es igual a la velocidad de los procesos que conducen a la destrucción del aducto.
La constante de Michaelis-Menten (KM) es una constante de equilibrio (referida al primer equilibrio descrito anteriormente); se puede decir, con una buena aproximación (porque también se debe considerar K3), que KM está representado por la relación entre las constantes cinéticas K2 y K1 (referidas a la destrucción y formación del aducto ES en el primer equilibrio descrito anteriormente) .
A través de la constante de Michaelis-Menten tenemos una "indicación de la afinidad entre enzima y sustrato: si la KM es pequeña c" es una "alta afinidad entre enzima y sustrato, entonces el aducto ES es estable.
Las enzimas están sujetas a regulación (o modulación).
En el pasado se hablaba principalmente de modulación negativa, es decir, inhibición de las capacidades catalíticas de una enzima, pero también puede haber una modulación positiva, es decir, existen especies capaces de potenciar las capacidades catalíticas de una enzima.
Hay 4 tipos de inhibiciones (obtenidas a partir de aproximaciones realizadas en un modelo para hacer coincidir los datos experimentales con ecuaciones matemáticas):
- inhibición competitiva
- inhibición no competitiva
- inhibición incompetitiva
- una inhibición competitiva
Hablamos de inhibición competitiva cuando una molécula (inhibidor) es capaz de competir con el sustrato. Por similitud estructural, el inhibidor puede reaccionar en lugar del sustrato, de ahí la terminología "inhibición competitiva" La probabilidad de que la enzima se una al inhibidor o sustrato depende de la concentración de ambos y de su afinidad con la enzima; por tanto, la velocidad de reacción depende de estos factores.
Para obtener la misma velocidad de reacción que sin la presencia del inhibidor, es necesario tener una mayor concentración de sustrato.
Se demuestra experimentalmente que, en presencia de un inhibidor, aumenta la constante de Michaelis-Menten.
En cuanto a, en cambio, la "inhibición no competitiva, la interacción entre la molécula que debería funcionar como moduladora (inhibidora positiva o negativa) y la" enzima, tiene lugar en un sitio diferente de aquel en el que la interacción ocurre entre la enzima y el sustrato; por tanto, hablamos de modulación alostérica (del griego alosteros → otro sitio).
Si el inhibidor se une a la enzima, puede inducir un cambio en la estructura de la enzima y, en consecuencia, puede disminuir la eficacia con la que el sustrato se une a la enzima.
En este tipo de procesos, la constante de Michaelis-Menten permanece constante ya que este valor depende de los equilibrios entre la enzima y el sustrato e, incluso en presencia de un inhibidor, estos equilibrios no cambian.
El fenómeno de la inhibición incompetitiva es raro; Un inhibidor incompetitivo típico es una sustancia que se une reversiblemente al aducto ES dando lugar a ESI:

La inhibición por exceso de sustrato a veces puede ser incompatible, ya que esto ocurre cuando una segunda molécula de sustrato se une al complejo ES, dando lugar al complejo ESS.
Un inhibidor competitivo, por otro lado, solo puede unirse al aducto enzimático del sustrato como en el caso anterior: la unión del sustrato a la enzima libre induce una modificación conformacional que hace que el sitio sea accesible para el inhibidor.
La constante de Michaelis Menten disminuye a medida que aumenta la concentración de inhibidor: aparentemente, por lo tanto, aumenta la afinidad de la enzima por el sustrato.
Serina proteasa
Son una familia de enzimas a las que pertenecen la quimotripsina y la tripsina.
La quimotripsina es una enzima proteolítica e hidrolítica que corta a la derecha de los aminoácidos hidrófobos y aromáticos.
El producto del gen que codifica la quimotripsina no está activo (se activa con un comando); la forma inactiva de quimotripsina está representada por una cadena polipeptídica de 245 aminoácidos. La quimotripsina tiene una forma globular debido a cinco puentes disulfuro y otras interacciones menores (electrostática, fuerzas de Van der Waals, enlaces de hidrógeno, etc.).
La quimotripsina es producida por las células quimosas del páncreas, donde está contenida en membranas especiales y expulsada a través del conducto pancreático hacia el intestino, en el momento de la digestión de los alimentos: la quimotripsina es de hecho una enzima digestiva. Las proteínas y nutrientes que ingerimos a través de la dieta se someten a digestión para reducirse a cadenas más pequeñas y ser absorbidas y transformadas en energía (por ejemplo, las amilasas y proteasas descomponen los nutrientes en glucosa y aminoácidos que llegan a las células, a través de los vasos sanguíneos. llegan a la vena porta y desde allí se conducen al hígado donde se someten a un tratamiento adicional).
Las enzimas se producen en forma inactiva y se activan solo cuando llegan al "sitio donde deben operar"; una vez finalizada su acción, se desactivan. Una enzima, una vez desactivada, no se puede reactivar: para tener una "acción catalítica adicional, debe ser reemplazada por" otra molécula de enzima. Si la chimitripsina se produjera en forma activa ya en el páncreas, atacaría a este último: las pancreatitis son patologías debidas a enzimas digestivas que ya están activadas en el páncreas (y no en los sitios requeridos); algunas de ellas si no se tratan a tiempo, conducir a la muerte.
En la quimotripsina y en todas las serina proteasas, la acción catalítica se debe a la existencia del anión alcohol (-CH2O-) en la cadena lateral de una serina.
Las serina proteasas toman este nombre precisamente porque su acción catalítica se debe a una serina.
Una vez que toda la enzima ha realizado su acción, antes de poder volver a operar sobre el sustrato nuevamente, se debe restaurar con agua; la "liberación" de serina por el agua es la etapa más lenta del proceso, y es esta fase. que determina la velocidad de catálisis.
La acción catalítica se produce en dos fases:
- formación del anión con propiedades catalíticas (alcoholato de anión) y posterior ataque nucleofílico sobre el carbono del carbonilo (C = O) con ruptura del enlace peptídico y formación del éster;
- Ataque de agua con restauración del catalizador (capaz de ejercer nuevamente su acción catalítica).
Las diversas enzimas pertenecientes a la familia de las serina proteasas pueden estar formadas por diferentes aminoácidos pero, para todos ellos, el sitio catalítico está representado por el anión alcohólico de la cadena lateral de una serina.
Una subfamilia de serina proteasas es la de las enzimas implicadas en la coagulación (que consiste en la transformación de la proteína, de su forma inactiva a una "otra forma que es activa). Estas enzimas aseguran que la coagulación sea lo más eficaz posible y esté limitada en el espacio y el tiempo (la coagulación debe ocurrir rápidamente y debe ocurrir solo en las proximidades del área lesionada) Las enzimas involucradas en la coagulación se activan en una cascada (a partir de la activación de una sola enzima, se obtienen miles de millones de enzimas: cada enzima activada , a su vez activa muchas otras enzimas).
La trombosis es una patología debida al mal funcionamiento de las enzimas de la coagulación: se produce por la activación, sin necesidad (porque no hay lesión), de las enzimas utilizadas en la coagulación.
Existen enzimas moduladoras (reguladoras) e inhibidoras de otras enzimas: interactuando con estas últimas, regulan o inhiben su actividad; incluso el producto de una enzima puede ser un inhibidor de la enzima.También hay enzimas que actúan cuanto más, mayor es el sustrato presente.
Lisozima
Luigi Pasteur descubrió, al estornudar en una placa de Petri, que en el moco hay una enzima capaz de matar bacterias: lisozima; del griego: liso = qué tamaño; zimo = enzima.
La lisozima es capaz de romper la pared celular de las bacterias. Las bacterias, y los organismos unicelulares en general, necesitan estructuras mecánicamente resistentes que limiten su forma; dentro de las bacterias hay una presión osmótica muy alta por lo que atraen el agua. La membrana plasmática explotaría si no hubiera una pared celular que se oponga a la entrada de agua y limite el volumen de la bacteria.
La pared celular consta de una cadena de polisacáridos en la que se alternan moléculas de N-acetil-glucosamina (NAG) y moléculas de ácido N-acetil-murámico (NAM); el enlace entre NAG y NAM se rompe por hidrólisis.El grupo carboxilo de NAM, en la pared celular, está comprometido en un enlace peptídico con un aminoácido.
Entre las diversas cadenas, se forman puentes que consisten en enlaces pseudo-péptidos: la ramificación se debe a la molécula de lisina; la estructura en su conjunto es muy ramificada y esto le confiere una alta estabilidad.
La lisozima es un antibiótico (mata las bacterias): actúa haciendo una grieta en la pared bacteriana; cuando esta estructura (que es resistente mecánicamente) se rompe, la bacteria extrae agua hasta que estalla. La lisozima logra romper el enlace glucosídico β-1,4 entre NAM y NAG.
El sitio catalítico de la lisozima está representado por un surco que corre a lo largo de la enzima en la que se inserta la cadena de polisacárido: seis anillos glucosídicos de la cadena se colocan en el surco.
En la posición tres de la ranura c "hay un estrangulador: en esta posición solo se puede colocar un NAG, porque el NAM, que es de mayores dimensiones, no puede entrar. El sitio catalítico real está entre las posiciones cuatro y cinco: ya que hay un NAG en la posición tres, el corte se realizará entre un NAM y un NAG (y no al revés); el corte, por tanto, es específico.
El pH óptimo para que la lisozima funcione es cinco. En el sitio catalítico de la enzima, es decir, entre las posiciones cuatro y cinco, se encuentran las cadenas laterales de un ácido aspártico y un ácido glutámico.
Grado de homología: mide el parentesco (es decir, similitud) entre estructuras de proteínas.
Existe una fuerte relación entre la lisozima y la lactosa sintasa.
La lactosa sintetasa sintetiza lactosa (que es el principal azúcar de la leche): la lactosa es un galactosil glucósido en el que c "es un enlace glucosídico β-1,4 entre la galactosa y la glucosa.
Por lo tanto, la lactosa sintetasa cataliza la reacción opuesta a la catalizada por la lisozima (que en su lugar divide el enlace glucosídico β-1,4)
La lactosa sintetasa es un dímero, es decir, está formado por dos cadenas de proteínas, una de las cuales tiene propiedades catalíticas y es comparable a la lisozima y la otra es una subunidad reguladora.
Durante el embarazo, las células de la glándula mamaria sintetizan glicoproteínas por la acción de la galatosiltransferasa (tiene una "homología de secuencia del 40% con la lisozima): esta enzima es capaz de transferir un grupo galactosilo de una estructura de alta energía a una estructura de glicoproteína. Durante el embarazo, se induce la expresión del gen que codifica la galactosisil-transferasa (también existe la expresión de otros genes que también dan otros productos): hay un aumento en el tamaño de la mama porque se activa la glándula mamaria (previamente inactiva) que debe producir leche Durante el parto se produce la α-lactoalbúmina que es una proteína reguladora: es capaz de regular la capacidad catalítica de la galactosil-transferasa (por discriminación del sustrato). La galactosil-transferasa modificada por α-lactalalbúmina es capaz de transferir un galactosilo a una molécula de glucosa: formando un enlace β-1,4 glicosídico y dando lactosa (lactosa sintetasa).
Por lo tanto, la galactosa transferasa prepara la glándula mamaria antes del parto y produce leche después del parto.
Para producir glicoproteínas, la galactosil transferasa se une a un galactosil y un NAG; durante el parto, la albúmina lactal se une a la galactosiltransferasa haciendo que esta última reconozca la glucosa y ya no NAG para dar lactosa.