" primera parte
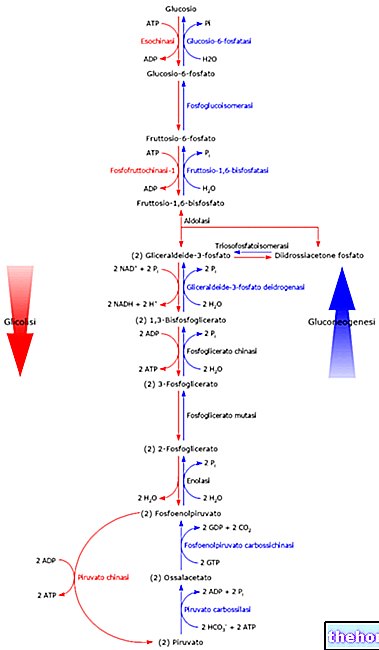
La enzima que cataliza la cuarta etapa del ciclo de krebs es la α-ceto glutarato deshidrogenasa; esta enzima es un complejo enzimático muy similar a la piruvato deshidrogenasa. Ambos están compuestos por 48-60 proteínas en las que se reconocen tres actividades enzimáticas diferentes y también tienen los mismos cofactores enzimáticos; son enzimas muy similares porque actúan sobre sustratos similares: tanto el piruvato como el l "α-ceto glutarato, son α- cetoácidos. El mecanismo de acción de los dos complejos enzimáticos es el mismo.
El ataque del pirofosfato de tiamina sobre el carbonilo (C = O) del "α-cetoglutarato, conduce a su descarboxilación y se forma el derivado carboxihidroxipropílico. Con la posterior transferencia a la lipoamida, tiene lugar un proceso redox interno, a partir del cual se obtiene el derivado carboxi de lipoamida o succinil lipoamida.
La succinil lipoamida luego reacciona con la coenzima A para dar succinil coenzima A (que continúa en el ciclo de krebs) y la lipoamida reducida que es reoxidada por el FAD: el FADH2 que se forma es reoxidado por NAD + y se obtiene NADH. En esta etapa, por tanto, tuvo lugar la segunda eliminación de un carbono del esqueleto carbonoso, en forma de dióxido de carbono.
Un grupo acilo unido a la coenzima A se encuentra en forma activada, es decir, tiene un alto contenido energético: por lo tanto, es posible aprovechar la energía de la succinil coenzima A.

En la quinta etapa del ciclo de krebs, la succinil coenzima A está sujeta a la acción de succinil tioquinasa; Se han formulado dos hipótesis sobre su modo de acción: describiremos solo una de las dos porque es la más acreditada. Según esta hipótesis, la succinil coenzima A es atacada por el nitrógeno de una histidina (Hys) de la enzima: se libera la coenzima A y se forma un aducto derivado de la histidina como intermedio, que es la succinil-enzima (o succinil-Hys ); un ortofosfato actúa sobre este intermedio, dando lugar a la liberación de succinato y la formación de la fosfoenzima. La fosfoenzima, atacada por el difosfato de guanosina (GDP), produce trifosfato de guasnosina (GTP) y se libera la enzima. Desde el punto de vista energético GTP = ATP: el enlace que aporta energía es el mismo en ambas especies (es el enlace anhídrido entre el fosforilo Β y el fosforilo γ). En algunos casos, el GTP se utiliza como material con un alto contenido energético pero, por lo general, el GTP se convierte en ATP por la acción de la enzima. nucleósido difosfo quinasa (NDPK); es una enzima que se encuentra en las células y cataliza la siguiente reacción:
N1TP + N2DP → N1DP + N2TP
Trifosfato de nucleósido genérico NiTP®
Difosfato de nucleósido genérico NiDP®
Es una reacción reversible; en nuestro caso sucede:
GTP + ADP → PIB + ATP
por lo tanto, puede avanzar hacia la derecha o hacia la izquierda incluso para pequeñas variaciones en las concentraciones de los reactivos.
Si el ciclo de krebs avanza a una velocidad tal que conduce a una producción de ATP superior a la energía requerida, hay escasa disponibilidad de ADP mientras que hay mucho ATP: la reacción catalizada por el nucleósido difosfo quinasa es, entonces, dirigido hacia la izquierda (GTP se acumula si el nucleósido difosfo quinasa no tiene suficiente sustrato, es decir, ADP). Por lo tanto, GTP es una señal de disponibilidad de energía y, por lo tanto, ralentiza el ciclo de krebs.
La sexta etapa del ciclo de Krebs conduce a la formación del fumarato por la acción de succinato deshidrogenasa; esta enzima da una reacción estereoespecífica ya que siempre se forma el trans insaturado (es un alqueno), es decir, el fumarato (mientras que el isómero cis es el maleato). La succinato deshidrogenasa se encuentra en la membrana mitocondrial interna, mientras que todas las demás enzimas del ciclo de krebs se encuentran dispersas por toda la mitocondria.
La succinato deshidrogenasa tiene FAD como cofactor; es inhibido por oxaloacetato (inhibición de retroalimentación) mientras que tiene succinato y fumarato como su modulador positivo (activador). su activador. Intentemos entender por qué, saltando a la etapa final del ciclo de krebs. La etapa final del ciclo de krebs. El ciclo de krebs requiere energía por lo que la única posibilidad de obtener oxaloacetato del paciente es que la concentración del paciente sea muy alta: el malato es uno de los metabolitos con mayor concentración en las células. La reacción que convierte el malato en oxaloacetato también se ve favorecida por el hecho de que la concentración de oxalacetato se mantiene baja por la acción de la citrato sintasa.La reacción catalizada por la succinato deshidrogenasa es, entonces, una reacción de autoalimentación y esta es la única forma de que se produzca la transformación del malato en oxalacetato.
La concentración de malato mitocondrial debe ser compatible con la concentración de malato citoplasmático: solo cuando la concentración de malato mitocondrial es tan alta como para garantizar la conversión del malato en oxaloacetato (en el ciclo de krebs), el malato también se puede utilizar en otras formas (que son citoplasmáticas): en el citoplasma el malato se puede convertir en oxaloacetato del que se puede obtener aspartato por acción de GOT (es una transaminasa) o glucosa por gluconeogénesis.
Volvemos a la séptima etapa del ciclo de krebs es catalizada por la enzima fumarasi: se añade agua de forma estereoespecífica para producir L-malato.
En la última etapa del ciclo de Krebs, de la que ya hemos hablado, la acción del malato deshidrogenasa. Esta enzima utiliza una molécula de NAD + para su acción catalítica.
Concluimos así la descripción de las distintas etapas del ciclo de krebs.
El ciclo de krebs es totalmente reversible.
Para aumentar la velocidad del ciclo de krebs, se puede aumentar la concentración de los metabolitos presentes en ese ciclo; Una de las estrategias para aumentar la velocidad del ciclo de krebs consiste en convertir parte del piruvato que ingresa a la mitocondria en oxaloacetato (por acción de la piruvato carboxilasa) y no transformarlo todo en acetil coenzima A: aumenta así la concentración de oxaloacetato que es un metabolito del ciclo de krebs y, por tanto, aumenta la velocidad de todo el ciclo.
En el ciclo de krebs se convierten tres NAD + en tres NADH y un FAD en FADH2 y, además, se obtiene un GTP: canalizando el poder reductor obtenido del ciclo de krebs, se produce más ATP; en la cadena respiratoria, el poder reductor se transfiere del NADH y FADH2 al oxígeno: esta transferencia se debe a una serie de enzimas ubicadas en la membrana mitocondrial que, en su acción, conducen a la producción de ATP.
Los procesos de la cadena respiratoria son procesos exergónicos y la energía liberada se utiliza para producir ATP; el propósito de la célula es explotar los procesos exergónicos para que se produzca la síntesis de ATP. Por cada molécula de NADH que ingresa a la cadena respiratoria, se obtienen 2.5 moléculas de ATP y por cada FADH2 se obtienen 1.5 moléculas de ATP; esta diversidad se debe al hecho de que FADH2 ingresa a la cadena respiratoria en un nivel más bajo que NADH.
Con el poder reductor del metabolismo aeróbico, se obtienen 30-32 ATP (219-233 kcal / mol) con una eficiencia de aproximadamente el 33% (la eficiencia del metabolismo anaeróbico es de aproximadamente el 2%).