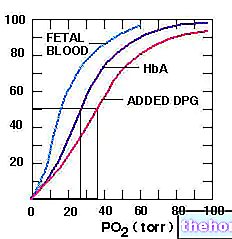
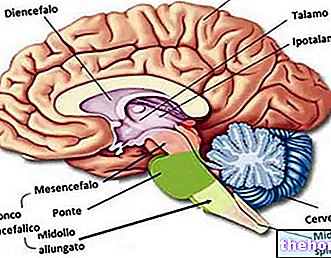
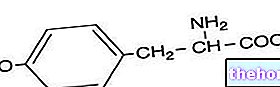
La síntesis de ácidos grasos comienza a partir de la acetil coenzima A y corresponde aproximadamente a la ruta inversa de su degradación; en la síntesis de ácidos grasos se añaden una serie de fragmentos de bicarbonato a la acetil coenzima A de partida.
La síntesis de ácidos grasos es completamente citoplasmática (es decir, las enzimas que catalizan esta síntesis se encuentran en el citoplasma). La acetil coenzima A utilizada en el citoplasma para la síntesis de ácidos grasos es de origen mitocondrial: una pequeña parte se transporta a través de la carnitina, por la acción de dos enzimas acil transferasas (una citoplasmática y otra mitocondrial) y una enzima translocasas. Parte de acetil La coenzima A de origen mitocondrial se obtiene a través de una ruta especializada: la citrato liasa (el nombre deriva de la primera enzima de este camino).
La acetil coenzima A presente en las mitocondrias se deriva de la glucólisis después de la acción de la piruvato deshidrogenasa; La acetil coenzima A sufre la acción de la enzima citrato sintasa: esta enzima cataliza la formación de citrato por reacción de acetil coenzima A con oxaloacetato.Si el ciclo de krebs es capaz de satisfacer las necesidades energéticas, parte del citrato (la cantidad innecesaria para el ciclo de krebs) puede salir de las mitocondrias y llegar al citoplasma, donde la enzima citrato liasa, gastando energía, la convierte nuevamente en acetil coenzima A y oxaloacetato De esta manera es posible tener acetil coenzima A disponible en el citoplasma, pero el oxalacetato que se forma debe ser devuelto a la mitocondria para estar disponible nuevamente para la enzima citrato sintasa.
El oxalacetato luego se transforma en malato por la acción de la enzima. malato deshidrogenasa citoplasmático (se gasta un NADH citoplásmico): el malato es un metabolito permeable y puede volver a entrar en la mitocondria donde, bajo la acción de la enzima malato deshidrogenasa mitocondrial, se reconvierte en oxaloacetato (también se obtiene un NADH); el paciente citoplasmático puede, alternativamente, someterse a la acción de la enzima málica, que realiza una descarboxilación y deshidrogenación, para convertirse en piruvato. La enzima málica actúa sobre NADP + (es similar al adenindinucleótido de nicotinamida pero, a diferencia de éste, tiene un grupo fosfórico en el segundo grupo hidroxilo en una de las dos unidades de ribosa) por lo tanto, en el paso del malato al piruvato, se produce NADPH ( que se usa en la biosíntesis) El piruvato ingresa a la mitocondria donde se transforma en oxaloacetato por la acción de la piruvato carboxilasa o en acetil coenzima A a través de la piruvato deshidrogenasa.
Veamos un ejemplo: se necesitan ocho moléculas de acetil coenzima A para sintetizar el ácido palmítico (cadena con dieciséis átomos de carbono) pero solo una de ellas se utiliza como tal: siete moléculas de acetil coenzima A son convertidas en malonil coenzima A por la enzima " acetil coenzima A carboxilasa (esta enzima usa una molécula de CO2 y tiene biotina como cofactor).
La enzima acetil coenzima A carboxilasa puede existir en una forma dispersa casi inactiva y en una forma agregada activa (alrededor de veinte unidades); la transición de la forma dispersa a la agregada ocurre cuando en el citoplasma hay una "alta concentración de citrato: el citrato es un modulador positivo de la enzima acetil coenzima A carboxilasa.
La enzima acetil coenzima A carboxilasa tiene otros moduladores positivos (insulina) y negativos (glucagón, adrenalina y acil coenzima A).
Analizaremos la síntesis de ácidos grasos en la bacteria escherichia coli en la que esta síntesis ocurre por la acción de siete proteínas distintas; en las células eucariotas, el mecanismo por el cual se produce la síntesis de ácidos grasos es similar al de las bacterias pero, en eucariotas, las siete enzimas responsables de la síntesis se agrupan en dos complejos multienzimáticos A y B.
En las bacterias, siete genes distintos codifican:
- ACP (proteína portadora de acilo);
- ACP-acetil transacetilasa;
- ACP.malonil transacetilasa;
- β-ceto-acil-ACP sintasa (enzima de condensación);
- β-ceto-acil-ACP reductasa;
- D-β-hidroxi-acil deshidratasa;
- enoil-ACP redactado.
En eucariotas, dos genes codifican:
Subunidad A
ACP;
Enzima condensadora
β-ceto-acil-ACP reductasa.
Subunidad B
ACP-acetil transacetilasa;
ACP-malonil transacetilasa;
D-β-hidroxi-acil deshidratasa;
enoil-ACP redactado.
Las siete proteínas de Escherichia coli están dispuestas de tal manera que hay una central (la ACP) y las otras seis a su alrededor.
En su acción enzimática intervienen dos grupos sulfhidrilo: uno que pertenece a una cisteína y otro que pertenece al brazo largo de una fosfopanteteína; El ACP se une al sustrato que, a través del brazo de la fosfopanteteína, se pone en contacto con las otras enzimas que, por tanto, pueden realizar su acción enzimática.
La acetil coenzima A (por medio de ACP acetil transacilasa) se une a la enzima ACP (más precisamente al azufre de la cisteína que forma el derivado de cisteína) y se libera la coenzima A; luego interviene la ACP-malonil transacilasa que cataliza el ataque de malonilo sobre fosfopanteteína (también en este proceso se libera la coenzima A que inicialmente estaba unida al malonilo).
El siguiente paso involucra la β-ceto-acil ACP sintasa que es una enzima de condensación: permite la fusión entre los dos esqueletos; el malonilo se descarboxila fácilmente y se forma un carbonilo del derivado de acetilo cisteína: se libera la cisteína y se forma un derivado de β-ceto (acetil acetil) fosfopantetina.
Posteriormente, interviene la β-ceto-acil-ACP reductasa que reduce aún más el carbonilo a la enzima ACP (se forma un hidróxido por NADPH que se reduce a NADP +).
Ahora, actúa la 3-hidroxi-acil ACP deshidratasa (se produce la deshidratación) lo que conduce a la formación de un sistema insaturado (alqueno).
El siguiente proceso involucra a la enoil-ACP-reductasa (realiza una hidrogenación: se forma el alcano y el NADPH se reduce a NADP +).
La última fase implica la conversión del producto acilo obtenido del primer ciclo en un compuesto capaz de iniciar un segundo ciclo: la enzima transacilasa transfiere el acilo a la cisteína, dejando libre el sitio de la pantetina que ahora estará dispuesta a unirse a otra. malonyl.
En la β-oxidación, se usa una molécula de FAD para obtener el metabolito α-β insaturado trans enoil coenzima A por deshidrogenación; en la síntesis de ácidos grasos, en cambio, se utiliza una molécula de NADPH para provocar la reacción opuesta.
Normalmente, se sintetizan ácidos grasos con dieciséis átomos de carbono, pero también se pueden producir ácidos grasos con dieciocho, veinte o veintidós átomos de carbono; los ácidos grasos se esterifican luego para formar triglicéridos con glicerol activado (es decir, glicerol 3-fosfato). Este último se puede obtener a partir de dihidroxiacetona fosfato por acción de la enzima glicerol fosfato deshidrogenasa o de glicerol a través de la enzima glicerol quinasa.
Los ácidos grasos sintetizados deben enviarse al tejido adiposo; se transportan en el torrente sanguíneo en forma de triglicéridos o, en parte como tales, con el uso de una proteína transportadora que es la albúmina.